Для всех микроорганизмов, входящих в царство, характерен прокариотический тип организации клетки, что определяется особенностями их ультраструктуры, а также строения и функций ряда макромолекул. Из всех известных клеток прокариотическая самая простая и, вероятно, первая клетка, возникшая около 3,6 млрд. лет назад.
В настоящее время предполагают, что в какой-то момент времени эволюция клеток пошла по двум самостоятельным направлениям. Появились две группы организмов -прокариоты, у которых ядерный материал не был ограничен оболочкой, и эукариоты, имеющие оформленное ядро с ядерной оболочкой.
Основные отличия прокариотов от эукариотов состоят в следующем:
в клетках прокариотов отсутствуют компартменты, или органеллы, ограниченные от цитоплазмы специализированными внутриклеточными липопротеидными мембранами: эндоплазматическая сеть (ретикулюм), митохондрии, аппарат Гольджи, лизосомы, хлоропласты;
ядерная структура прокариотов, называемая нуклеоидом, не имеет ядерной оболочки с поровым комплексом и представлена макромолекулой ДНК с белками (без гистонов);
геном прокариотической клетки организован в одну кольцевую хромосому, которая представляет собой единый репликон и не делится митозом;
дополнительные репликоны могут быть представлены кольцевыми молекулами плазмидных ДНК;
прокариотическая клетка содержит только один тип рибосом с константой седиментации 708, причем часть рибосом ассоциирована с цитоплазматической мембраной, что никогда не наблюдается у эукариотов;
клеточная стенка прокариотов содержит характерный только для бактерий биогетерополимер - пептидогликан.
Некоторые прокариоты обладают структурами, отсутствующими у эукариотов:
подвижные бактерии имеют особые бактериальные жгутики из белков-флагеллинов;
спорообразующие формы бактерий в неблагоприятных условиях превращаются в уникальные по степени устойчивости типы покоящихся клеток - бактериальные споры;
клетки прокариотов очень малы; диаметр большинства клеток бактерий не превышает 1 мкм, однако длина может быть значительной, например, у некоторых спирохет - до 500 мкм. Малые размеры прокариотов связаны, как полагают, с отсутствием в их ультраструктуре специализированных мембранных систем, что затрудняет координацию внутриклеточных процессов пропорционально увеличению размеров клетки.
Клеточное строение четко отделяет прокариотов от вирусов. Подчеркивая примитивность организации бактериальных клеток, необходимо однако отметить, что они эволюционировали в своем направлении в течение гораздо большего времени, чем эукарио-тические, и, хотя эволюционные возможности прокариотической клетки, по всей видимости, ограничены, в процессе эволюции происходили изменения их клеточной организации, что привело постепенно к ее усложнению.
По целому ряду признаков бактерии имеют принципиальные различия с эукариотами, и знание особенностей их устройства и функционирования позволяет понять возможность избирательного антимикробного действия химиотерапевтических препаратов. Применение электронной микроскопии и тонких цитохимических исследований позволило изучить их ультраструктуру (рис. 1). Обязательными компонентами бактериальной клетки являются цитоплазматическая мембрана, окружающая цитоплазму, в которой содержатся рибосомы и нуклеоид. Клетки всех бактерий, за исключением L-форм и микоплазм, имеют клеточную стенку. Другие структуры являются дополнительными и определяют морфологические и функциональные особенности различных видов: капсулы, жгутики, пили, споры, включения.
Рис. 1. Схема строения прокариотической клетки:
/ - капсула; 2 - клеточная стенка; 3 - цитоплазматическая мембрана; 4 - мезосомы; 5 - цитоплазма; 6 - нуклеоид; 7 - плазмида;
8 - рибосомы и полисомы; 9 - жгутики; 10 - пили; 11 - гранулы гликогена; 12 - липидные капельки; 13 - гранулы волютина; 14 - включения серы
Поверхностные структуры. Капсула - это наружный, самый верхний слизистый слой клетки различной толщины фибриллярной или глобулярной структуры. Она имеет полисахаридную, му-кополисахаридную или полипептидную природу и содержит до 98 % воды. В зависимости от толщины различают микрокапсулу (толщиной менее 0,2 мкм) и макрокапсулу. Капсула не является обязательным структурным элементом клетки. Биологический смысл капсулообразования определяется целым рядом функций, среди которых: защита от фагоцитов и вирусов, токсинов и радиации; иммунологическая мимикрия у патогенных бактерий; сохранение влаги в условиях пониженной влажности; прикрепление клетки к плотной поверхности.
Пили (фимбрии, ворсинки, реснички) - это прямые цилиндрические образования белковой природы длиной 0,3-10 мкм, диаметром до 10 нм, равномерно покрывающие поверхность клетки (до нескольких сотен на клетку), не выполняющие локомоторную функцию.
Различают пили общего типа, способствующие прикреплению бактериальной клетки к субстрату, клеткам человека (явление адгезии микроорганизмов) и пили половые, участвующие в передаче генетического материала от клетки-донора к клетке-реципиенту в процессе конъюгации, а также обуславливающие адсорбцию специфических бактериофагов на клетках.
Жгутики - органы движения бактерий в виде спирально изогнутых цилиндрических образований белковой природы (белки-флагеллины) на поверхности клетки длиной 3-12 мкм и толщиной 10-30 нм, прикрепленные базальным телом (системой дисков) к цитоплазматической мембране (см. вкл. I). Число и располо-
жение жгутиков может быть различным и является видовым признаком (рис. 2). Различают монотрихи (бактерии с одним жгутиком на конце), амфитрихи (бактерии с жгутиками, расположенными по полюсам), лофотрихи (клетки с пучком жгутиков на одном конце) и перитрихи (с 2-30 жгутиками по всему телу клетки).
Пили и жгутики не являются обязательными органоидами бактериальной клетки.
Клеточная стенка - один из основных структурных элементов бактерии, выполняющий механическую защиту клетки. Кроме микоплазм и L-форм, клетки всех бактерий покрыты клеточной стенкой, толщина которой у разных видов колеблется в пределах 0,01-14 мкм. Она представляет собой плотную эластичную структу -
Рис. 2. Основные формы бактерий (по А. А. Воробьеву и соавт., 1994):
/ - стафилококки; 2 - стрептококки; 3 - сарцины; 4 - гонококки;
5- пневмококки; 6- капсулы пневмококков; 7- коринебактерии дифтерии; 8 - клостридии; 9 - бациллы; 10 - вибрионы; 11 - спириллы; 12 - трепонсмы; - боррелии; 14 - лептоспиры; 15- актиномицеты; 16 - расположение жгутиков: а - монотрихи; б - лофотрихи; в - амфитрихи; г - перитрихи
ру, которая окружает протопласт клетки и придает ей постоянную форму и жесткость. Клеточная стенка препятствует осмотическому набуханию и разрыву клеток, когда они попадают в гипотоническую среду. Вода, другие малые молекулы и разные ионы легко проникают через крошечные поры в клеточной стенке, но через них не проходят крупные молекулы белков и нуклеиновых кислот.
Основным химическим компонентом клеточной стенки является специфический гетерополимер - пептидогликан (муреин, мукопептид, глюкозаминопептид, гликопептид), состоящий из цепочек, в которых чередуются остатки N-ацетилглюкозамина и М-ацетилмурамовой кислоты, соединенные между собой β-1,4-гликозидными связями. Это резко отличает оболочечные структуры бактерий от эукариотических и создает «ахиллесову пяту» бактерий, используемую для антимикробной химиотерапии.
Организация цитоплазмы. Цитоплазматическая мембрана (ЦМ) Носится к числу обязательных клеточных структур, имеет толщину 7-13 нм и располагается непосредственно под клеточной стенкой, ограничивая протопласт клетки. По своему строению мембраны бактериальных, животных и растительных клеток очень сходны. В настоящее время большинством ученых принята жидкостно-мозаичная модель строения ЦМ. Согласно этой модели ЦМ состоит из двойного слоя (15-30 % молекул фосфолипидов и триглицеридов с направленными внутрь гидрофобными концами и гидрофильными «головками» наружу. В него мозаично погружены молекулы белка (50-70%). В мембране имеются также углеводы (2-5 %) и РНК. ЦМ представляет собой пластическое «текучее» образование, которое играет важнейшую роль обмене веществ, является полупроницаемой структурой, поддерживает осмотическое давление, контролирует как поступление веществ в клетку, так и выведение конечных метаболитов по системе субстрат специфичных пермеаз (ферментов-переносчиков, локализованных на мембране). С ЦМ связаны процессы дыхания, доставляющие клетке энергию, то есть те функции, за которые в эукариотической клетке ответственны мембраны митохондрий и хлоропластов.
Выделяют так называемые мезосомы - впячивания ЦМ - смешанные мембранные системы, образованные трубочками, пузырьками и ламеллами. Предполагается выполнение ими функций центра дыхательной активности бактерий, участие в делении клетки и расхождении дочерних хромосом после репликации.
Цитоплазма заполняет собой объем бактерии, ограниченный ЦМ. Это сложная коллоидная система, которая состоит из бельков, нуклеиновых кислот, углеводов, липидов, минеральных веществ и 70-80 % воды. Цитоплазма является местонахождением внутриклеточных органелл (нуклеоида, рибосом, различных включений) и участвует во внутриклеточном метаболизме. Характер-
ными чертами организации цитоплазмы прокариотов по сравнению с животными и растительными клетками являются отсутствие эндоплазматического ретикулюма и высокая электронная плотность.
Нуклеоид - ядерный материал бактериальной клетки. Он представлен двойной нитью макромолекулы ДНК с молекулярной массой 2-3 10 в соединении с белками, среди которых отсутствуют характерные для эукариотов ядерные (гистоны и гистоноподобные) белки. В отличие от настоящего ядра эукариотических клеток нуклеоид не имеет ядерной перфорированной мембраны, не делится митозом и представляет собой в период деления одну кольцевую хромосому, кодирующую всю генетическую информацию.
Плазмиды - необязательные внутриклеточные структуры в виде внехромосомных кольцевых участков ДНК, способных к саморепликации. Обуславливают наследование дополнительных признаков: лекарственной устойчивости, токсигенности, бактериоциногенности и др.
Рибосомы - органеллы, в которых осуществляется синтез белка. Каждая рибосома имеет размеры 20х30х30 нм и константу седиментации 70S (так как при ультрацентрифугировании рибосомы оседают со скоростью около 70 единиц Сведенберга (S), в отличие от более крупных цитоплазматических рибосом эукариотов с константой седиментации 808). В свободном состоянии бактериальная рибосома находится в виде двух субъединиц - 30S и 50S, обе субъединицы содержат примерно по 40 % рибосомальной РНК и 60 % белка. Во время синтеза белка рибосомы с помощью информационной РНК образуют полисомы, обычно связанные с ЦМ. Бактерии могут содержать от 5000 до 50 000 рибосом, что зависит от возраста клетки и условий культивирования.
Знание различий между рибосомами бактерий и эукариотических клеток имеет важное значение для понимания механизмов антимикробного действия тех антибиотиков, которые подавляют синтез белка на бактериальных рибосомах и не затрагивают функции 80S рибосом.
Споры (эндоспоры) бактерий - покоящиеся формы некоторых видов грамположительных бактерий в неблагоприятных условиях внешней среды.
Спорообразование происходит в несколько стадий, при полном созревании споры вегетативная часть клетки лизируется и отмирает (см. вкл. I, II).
В процессе спорообразования (споруляции) можно выделить несколько основных этапов. Переходящая к спорообразованию клетка перестает расти; как правило, она содержит два и более нуклеоидов. На первом этапе часть клеточной ДНК локализуется в одном из полюсов клетки. Затем часть цитоплазмы с заключен-
Иной в ней хромосомой обособляется цитоплазматической мембраной, как бы врастающей в глубь клетки, при этом образуется проспорь, окруженная двойной мембранной оболочкой.
Затем между двумя мембранами идет формирование многослойной стенки и коры (кортекса) споры пептидогликановой природы. Снаружи мембран образуется также полипептидная оболочка и экзоспорий, окружающий спору в виде свободного чехла. Полностью сформировавшаяся бактериальная спора - это уплотненный участок клетки с нуклеоидом и рибосомами, ограниченный плотной многослойной оболочкой, пропитанной кальциевыми Солями дипиколиновой кислоты.
Спорообразование характерно для палочковидных бактерий - бацилл и клостридий (см. рис. 2). Различают центральное, терминальное и субтерминальное расположение спор в вегетативной части клетки, что является дифференциально-диагностическим признаком возбудителя.
В одной бактерии образуется одна спора, находящаяся в стадии покоя, при этом все процессы обмена веществ практически сведены к нулю, но сохраняется потенциальная жизнеспособность клетки. Поскольку увеличение числа микроорганизмов в этом процессе не происходит, спорообразование у бактерий не является способом размножения, а лишь приспособлением для выживания. Уникальные по степени своей устойчивости к физическим и химическим факторам бактериальные споры могут сохраняться до внешней среде без потери жизнеспособности длительное время (десятки лет), затрудняя борьбу со спороносными патогенными бактериями.
Внутриплазматические включения. Термином «включения» обозначают такие внутриклеточные структуры бактерий, которые, очевидно, не являются абсолютно необходимыми для их жизнедеятельности. Однако их природа и функции могут быть различны. В одних случаях включения являются продуктами обмена бактериальной клетки, в других - запасом питательных веществ.
Из резервных полисахаридов особенно распространены глюканы - гликоген, крахмал, гранулеза. Они выявляются в клетках бацилл, клостридий, энтеробактерий и др.
Запасные липиды представлены полиэфиром β-оксимасляной кислоты и восками. Воски, эфиры высокомолекулярных жирных кислот и спиртов характерны для микобактерий.
У коринебактерий резерв фосфора создается в виде зерен полифосфатов (волютина), имеющих диагностическое значение.
К прокариотам относятся архебактерии, бактерии и синезеленые водоросли. Прокариоты — одноклеточные организмы, у которых отсутствуют структурно оформленное ядро, мембранные органоиды и митоз.
Размеры — от 1 до 15 мкм. Основные формы: 1) кокки (шаровидные), 2) бациллы (палочковидные), 3) вибрионы (изогнутые в виде запятой), 4) спириллы и спирохеты (спирально закрученные).
1 — кокки; 2 — бациллы; 3 — вибрионы; 4—7 — спириллы и спирохеты.

1 — цитоплазматическая мемб-рана; 2 — клеточ-ная стенка; 3 — слизис-тая кап-сула; 4 — цито-плазма; 5 — хромо-сомная ДНК; 6 — рибосомы; 7 — мезо-сома; 8 — фото-синтети-ческие мемб-раны; 9 — вклю-чения; 10 — жгу-тики; 11 — пили.
Бактериальная клетка ограничена оболочкой. Внутренний слой оболочки представлен цитоплазматической мембраной (1), над которой находится клеточная стенка (2); над клеточной стенкой у многих бактерий — слизистая капсула (3). Строение и функции цитоплазматической мембраны эукариотической и прокариотической клеток не отличаются. Мембрана может образовывать складки, называемые мезосомами (7). Они могут иметь разную форму (мешковидные, трубчатые, пластинчатые и др.).
На поверхности мезосом располагаются ферменты. Клеточная стенка толстая, плотная, жесткая, состоит из муреина (главный компонент) и других органических веществ. Муреин представляет собой правильную сеть из параллельных полисахаридных цепей, сшитых друг с другом короткими белковыми цепочками. В зависимости от особенностей строения клеточной стенки бактерии подразделяются на грамположительные (окрашиваются по Граму) и грамотрицательные (не окрашиваются). У грамотрицательных бактерий стенка тоньше, устроена сложнее и над муреиновым слоем снаружи имеется слой липидов. Внутреннее пространство заполнено цитоплазмой (4).
Генетический материал представлен кольцевыми молекулами ДНК. Эти ДНК можно условно разделить на «хромосомные» и плазмидные. «Хромосомная» ДНК (5) — одна, прикреплена к мембране, содержит несколько тысяч генов, в отличие от хромосомных ДНК эукариот она не линейная, не связана с белками. Зона, в которой расположена эта ДНК, называется нуклеоидом . Плазмиды — внехромосомные генетические элементы. Представляют собой небольшие кольцевые ДНК, не связаны с белками, не прикреплены к мембране, содержат небольшое число генов. Количество плазмид может быть различным. Наиболее изучены плазмиды, несущие информацию об устойчивости к лекарственным препаратам (R-фактор), принимающие участие в половом процессе (F-фактор). Плазмида, способная объединяться с хромосомой, называется эписомой .
В бактериальной клетке отсутствуют все мембранные органоиды, характерные для эукариотической клетки (митохондрии, пластиды, ЭПС, аппарат Гольджи, лизосомы).
В цитоплазме бактерий находятся рибосомы 70S-типа (6) и включения (9). Как правило, рибосомы собраны в полисомы. Каждая рибосома состоит из малой (30S) и большой субъединиц (50S). Функция рибосом: сборка полипептидной цепочки. Включения могут быть представлены глыбками крахмала, гликогена, волютина, липидными каплями.
У многих бактерий имеются жгутики (10) и пили (фимбрии) (11). Жгутики не ограничены мембраной, имеют волнистую форму и состоят из сферических субъединиц белка флагеллина. Эти субъединицы расположены по спирали и образуют полый цилиндр диаметром 10-20 нм. Жгутик прокариот по своей структуре напоминает одну из микротрубочек эукариотического жгутика. Количество и расположение жгутиков может быть различным. Пили — прямые нитевидные структуры на поверхности бактерий. Они тоньше и короче жгутиков. Представляют собой короткие полые цилиндры из белка пилина. Пили служат для прикрепления бактерий к субстрату и друг к другу. Во время конъюгации образуются особые F-пили, по которым осуществляется передача генетического материала от одной бактериальной клетки к другой.
Спорообразование у бактерий — способ переживания неблагоприятных условий. Споры формируются обычно по одной внутри «материнской клетки» и называются эндоспорами. Споры обладают высокой устойчивостью к радиации, экстремальным температурам, высушиванию и другим факторам, вызывающим гибель вегетативных клеток.
Размножение. Бактерии размножаются бесполым способом — делением «материнской клетки» надвое. Перед делением происходит репликация ДНК.
Редко у бактерий наблюдается половой процесс, при котором происходит рекомбинация генетического материала. Следует подчеркнуть, что у бактерий никогда не образуются гаметы, не происходит слияние содержимого клеток, а имеет место передача ДНК от клетки-донора к клетке-реципиенту. Различают три способа передачи ДНК: конъюгация, трансформация, трансдукция.

— однонаправленный перенос F-плазмиды от клетки-донора в клетку-реципиента, контактирующих друг с другом. При этом бактерии соединяются друг с другом особыми F-пилями (F-фимбриями), по каналам которых фрагменты ДНК и переносятся. Конъюгацию можно разбить на следующие этапы: 1) раскручивание F-плазмиды, 2) проникновение одной из цепей F-плазмиды в клетку-реципиента через F-пилю, 3) синтез комплементарной цепи на матрице одноцепочечной ДНК (происходит как в клетке-доноре (F +), так и в клетке-реципиенте (F -)).
Трансформация — однонаправленный перенос фрагментов ДНК от клетки-донора к клетке-реципиенту, не контактирующих друг с другом. При этом клетка-донор или «выделяет» из себя небольшой фрагмент ДНК, или ДНК попадает в окружающую среду после гибели этой клетки. В любом случае ДНК активно поглощается клеткой-реципиентом и встраивается в собственную «хромосому».
Трансдукция — перенос фрагмента ДНК от клетки-донора к клетке-реципиенту с помощью бактериофагов.
Вирусы
Вирусы состоят из нуклеиновой кислоты (ДНК или РНК) и белков, образующих оболочку вокруг этой нуклеиновой кислоты, т.е. представляют собой нуклеопротеидный комплекс. В состав некоторых вирусов входят липиды и углеводы. Вирусы содержат всегда один тип нуклеиновой кислоты — либо ДНК, либо РНК. Причем каждая из нуклеиновых кислот может быть как одноцепочечной, так и двухцепочечной, как линейной, так и кольцевой.
Размеры вирусов — 10-300 нм. Форма вирусов: шаровидная, палочковидная, нитевидная, цилиндрическая и др.
Капсид — оболочка вируса, образована белковыми субъединицами, уложенными определенным образом. Капсид защищает нуклеиновую кислоту вируса от различных воздействий, обеспечивает осаждение вируса на поверхности клетки-хозяина. Суперкапсид характерен для сложноорганизованных вирусов (ВИЧ, вирусы гриппа, герпеса). Возникает во время выхода вируса из клетки-хозяина и представляет собой модифицированный участок ядерной или наружной цитоплазматической мембраны клетки-хозяина.
Если вирус находится внутри клетки-хозяина, то он существует в форме нуклеиновой кислоты. Если вирус находится вне клетки-хозяина, то он представляет собой нуклеопротеидный комплекс, и эта свободная форма существования называется вирионом . Вирусы обладают высокой специфичностью, т.е. они могут использовать для своей жизнедеятельности строго определенный круг хозяев.
В цикле репродукции вируса можно выделить следующие стадии.

- Осаждение на поверхности клетки-хозяина.
- Проникновение вируса в клетку-хозяина (могут попасть в клетку-хозяина путем: а) «инъекции», б) растворения оболочки клетки вирусными ферментами, в) эндоцитоза; попав внутрь клетки вирус переводит ее белок-синтезирующий аппарат под собственный контроль).
- Встраивание вирусной ДНК в ДНК клетки-хозяина (у РНК-содержащих вирусов перед этим происходит обратная транскрипция — синтез ДНК на матрице РНК).
- Транскрипция вирусной РНК.
- Синтез вирусных белков.
- Синтез вирусных нуклеиновых кислот.
- Самосборка и выход из клетки дочерних вирусов. Затем клетка либо погибает, либо продолжает существовать и производить новые поколения вирусных частиц.

Вирус иммунодефицита человека поражает главным образом CD 4 -лимфоциты (хелперы), на поверхности которых есть рецепторы, способные связываться с поверхностным белком ВИЧ. Кроме того, ВИЧ проникает в клетки ЦНС, нейроглии, кишечника. Иммунная система организма человека утрачивает свои защитные свойства и оказывается не в состоянии противостоять возбудителям различных инфекций. Средняя продолжительность жизни инфицированного человека составляет 7-10 лет.
Источником заражения служит только человек — носитель вируса иммунодефицита. СПИД передается половым путем, через кровь и ткани, содержащие вирус иммунодефицита, от матери к плоду.
Перейти к лекции №8 « Ядро. Хромосомы»
Перейти к лекции №10 « Понятие об обмене веществ. Биосинтез белков»
К прокариотам относят бактерии и сине-зелёные водоросли (цианеи). Наследственный аппарат прокариот представлен одной кольцевой молекулой ДНК, не образующей связей с белками и содержащей по одной копии каждого гена - гаплоидные организмы. В цитоплазме имеется большое количество мелких рибосом; отсутствуют или слабо выражены внутренние мембраны. Ферменты пластического обмена расположены диффузно. Аппарат Гольджи представлен отдельными пузырьками. Ферментные системы энергетического обмена упорядоченно расположены на внутренней поверхности наружной цитоплазматической мембраны. Снаружи клетка окружена толстой клеточной стенкой. Многие прокариоты способны к спорообразованию в неблагоприятных условиях существования; при этом выделяется небольшой участок цитоплазмы содержащий ДНК, и окружается толстой многослойной капсулой. Процессы метаболизма внутри споры практически прекращаются. Попадая в благоприятные условия, спора преобразуется в активную клеточную форму. Размножение прокариот происходит простым делением надвое.
Прокариотические и эукариотические клетки (Т.А. Козлова, В.С. Кучменко. Биология в таблицах. М.,2000)
| Признаки | Прокариоты | Эукариоты |
| 1 ЯДЕРНАЯ МЕМБРАНА | Отсутствует | Имеется |
| ПЛАЗМАТИЧЕСКАЯ МЕМБРАНА | Имеется | Имеется |
| МИТОХОНДРИИ | Отсутствуют | Имеются |
| ЭПС | Отсутствует | Имеется |
| РИБОСОМЫ | Имеются | Имеются |
| ВАКУОЛИ | Отсутствуют | Имеются (особенно характерны для растений) |
| ЛИЗОСОМЫ | Отсутствуют | Имеются |
| КЛЕТОЧНАЯ СТЕНКА | Имеется, состоит из сложного гетерополимерного вещества | Отсутствует в животных клетках, в растительных состоит из целлюлозы |
| КАПСУЛА | Если имеется, то состоит из соединений белка и сахара | Отсутствует |
| КОМПЛЕКС ГОЛЬДЖИ | Отсутствует | Имеется |
| ДЕЛЕНИЕ | Простое | Митоз, амитоз, мейоз |
Другие записи
10.06.2016. Химическая организация клетки. Неорганические вещества
Изучение химического состава клеток показывает, что в живых организмах нет никаких особых химических элементов, свойственных только им: именно в этом проявляется единство химического состава живой и…
10.06.2016. Строение эукариотической клетки
Клетки, образующие ткани животных и растений, значительно различаются по форме, размерам и внутреннему строению. Однако все они обнаруживают сходство в главных чертах процессов жизнедеятельности, обмена…
Тип урока : комбинированный.
Методы : словесный, наглядный, практический, проблемно-поисковый.
Цели урока
Образовательная: углубить знания учащихся о строении клеток эукариот, научить применять их на практических занятиях.
Развивающие: совершенствовать умения учащихся работать с дидактическим материалом; развивать мышление учащихся, предлагая задания для сравнения клеток прокариот и эукариот, клеток растений и клетки животных с выявлением схожих и отличительных признаков.
Оборудование : плакат «Строение цитоплазматической мембраны»; карточки-задания; раздаточный материал (строение прокариотической клетки, типичная растительная клетка, строение животной клетки).
Межпредметные связи : ботаника, зоология, анатомия и физиология человека.
План урока
I. Организационный момент
Проверка готовности к уроку.
Проверка списочного состава учащихся.
Сообщение темы и целей урока.
II. Изучение нового материала
Разделение организмов на про- и эукариоты
По форме клетки необычайно разнообразны: одни имеют округлую форму, другие похожи на звездочки со многими лучами, третьи вытянутые и т.д. Различны клетки и по размеру – от мельчайших, с трудом различимых в световом микроскопе, до прекрасно видимых невооруженным глазом (например, икринки рыб и лягушек).
Любое неоплодотворенное яйцо, в том числе гигантские окаменевшие яйца ископаемых динозавров, которые хранятся в палеонтологических музеях, тоже были когда-то живыми клетками. Однако, если говорить о главных элементах внутреннего строения, все клетки схожи между собой.
Прокариоты (от лат. pro – перед, раньше, вместо и греч. karyon – ядро) – это организмы, клетки которых не имеют ограниченного мембраной ядра, т.е. все бактерии, включая архебактерии и цианобактерии. Общее число видов прокариот около 6000. Вся генетическая информация прокариотической клетки (генофор) содержится в одной-единственной кольцевой молекуле ДНК. Митохондрии и хлоропласты отсутствуют, а функции дыхания или фотосинтеза, обеспечивающие клетку энергией, выполняет плазматическая мембрана (рис. 1). Размножаются прокариоты без выраженного полового процесса путем деления надвое. Прокариоты способны осуществлять целый ряд специфических физиологических процессов: фиксируют молекулярный азот, осуществляют молочнокислое брожение, разлагают древесину, окисляют серу и железо.
После вступительной беседы учащиеся рассматривают строение прокариотической клетки, сравнивая основные особенности строения с типами эукариотической клетки (рис. 1).
Эукариоты – это высшие организмы, имеющие четко оформленное ядро, которое оболочкой отделяется от цитоплазмы (кариомембраной). К эукариотам относятся все высшие животные и растения, а также одноклеточные и многоклеточные водоросли, грибы и простейшие. Ядерная ДНК у эукариот заключена в хромосомах. Эукариоты обладают клеточными органоидами, ограниченными мембранами.
Отличия эукариот от прокариот
– Эукариоты имеют настоящее ядро: генетический аппарат эукариотической клетки защищен оболочкой, схожей с оболочкой самой клетки.
– Включенные в цитоплазму органоиды окружены мембраной.
Строение клеток растений и животных
Клетка любого организма представляет собой сис-тему. Она состоит из трех взаимосвязанных между собой частей: оболочки, ядра и цитоплазмы.
При изучении ботаники, зоологии и анатомии человека вы уже знакомились со строением различных типов клеток. Кратко повторим этот материал.
Задание 1. Определите по рисунку 2, каким организмам и типам тканей соответствуют клетки под цифрами 1–12. Чем обусловлена их форма?
Строение и функции органоидов растительных и животных клеток
Используя рисунки 3 и 4 и пользуясь Биологическим энциклопедическим словарем и учебником, учащиеся заполняют таблицу, сравнивая животную и растительную клетки.
Таблица. Строение и функции органоидов растительных и животных клеток
Органоиды клетки |
Строение органоидов |
Функция |
Присутствие органоидов в клетках |
|
растений |
животных |
|||
Хлоропласт |
Представляет собой разновидность пластид |
Окрашивает растения в зеленый цвет, в нем происходит фотосинтез |
||
Лейкопласт |
Оболочка состоит из двух элементарных мембран; внутренняя, врастая в строму, образует немногочисленные тилакоиды |
Синтезирует и накапливает крахмал, масла, белки |
||
Хромопласт |
Пластиды с желтой, оранжевой и красной окраской, окраска обусловлена пигментами – каротиноидами |
Красная, желтая окраска осенних листьев, сочных плодов и др. |
||
Занимает до 90% объема зрелой клетки, заполнена клеточным соком |
Поддержание тургора, накопление запасных веществ и продуктов обмена, регуляция осмотического давления и др. |
|||
Микротрубочки |
Состоят из белка тубулина, расположены около плазматической мембраны |
Участвуют в отложении целлюлозы на клеточных стенках, перемещении в цитоплазме различных органоидов. При делении клетки микротрубочки составляют основу структуры веретена деления |
||
Плазматическая мембрана (ЦПМ) |
Состоит из липидного бислоя, пронизанного белками, погруженными на различную глубину |
Барьер, транспорт веществ, сообщение клеток между собой |
||
Гладкий ЭПР |
Система плоских и ветвящихся трубочек |
Осуществляет синтез и выделение липидов |
||
Шероховатый ЭПР |
Название получил из-за множества рибосом, находящихся на его поверхности |
Синтез белков, их накопление и преобразование для выделения из клетки наружу |
||
Окружено двойной ядерной мембраной, имеющей поры. Наружная ядерная мембрана образует непрерывную структуру с мембраной ЭПР. Содержит одно или несколько ядрышек |
Носитель наследственной информации, центр регуляции активности клетки |
|||
Клеточная стенка |
Состоит из длинных молекул целлюлозы, собранных в пучки, называемые микрофибриллами |
Внешний каркас, защитная оболочка |
||
Плазмодесмы |
Мельчайшие цитоплазматические каналы, которые пронизывают клеточные стенки |
Объединяют протопласты соседних клеток |
||
Митохондрии |
Синтез АТФ (аккумуляция энергии) |
|||
Аппарат Гольджи |
Состоит из стопки плоских мешочков – цистерн, или диктиосом |
Синтез полисахаридов, формирование ЦПМ и лизосом |
||
Лизосомы |
Внутриклеточное пищеварение |
|||
Рибосомы |
Состоят из двух неравных субъединиц – |
Место биосинтеза белка |
||
Цитоплазма |
Состоит из воды с большим количеством растворенных в ней веществ, содержащих глюкозу, белки и ионы |
В ней расположены другие органоиды клетки и осуществляются все процессы клеточного метаболизма |
||
Микрофиламенты |
Волокна из белка актина, обычно располагаются пучками вблизи поверхности клеток |
Участвуют в подвижности и изменении формы клеток |
||
Центриоли |
Могут входить в состав митотического аппарата клетки. В диплоидной клетке содержится две пары центриолей |
Участвуют в процессе деления клетки у животных; в зооспорах водорослей, мхов и у простейших образуют базальные тельца ресничек |
||
Микроворсинки |
Выступы плазматической мембраны |
Увеличивают наружную поверхность клетки, микроворсинки в совокупности образуют кайму клетки |
||
Выводы
1. Клеточная стенка, пластиды и центральная вакуоль присущи только растительным клеткам.
2. Лизосомы, центриоли, микроворсинки присутствуют в основном только в клетках животных организмов.
3. Все остальные органоиды характерны как для растительных, так и для животных клеток.
Строение оболочки клеток
Клеточная оболочка располагается снаружи клетки, отграничивая последнюю от внешней или внутренней среды организма. Ее основу составляет плазмалемма (клеточная мембрана) и углеводно-белковая составляющая.
Функции клеточной оболочки:
– поддерживает форму клетки и придает механическую прочность клетке и организму в целом;
– защищает клетку от механических повреждений и попадания в нее вредных соединений;
– осуществляет узнавание молекулярных сигналов;
– регулирует обмен веществ между клеткой и средой;
– осуществляет межклеточное взаимодействие в многоклеточном организме.
Функция клеточной стенки:
– представляет собой внешний каркас – защитную оболочку;
– обеспечивает транспорт веществ (через клеточную стенку проходит вода, соли, молекулы многих органических веществ).
Наружный слой клеток животных, в отличие от клеточных стенок растений, очень тонкий, эластичный. Он не виден в световой микроскоп и состоит из разнообразных полисахаридов и белков. Поверхностный слой животных клеток называется гликокаликсом , выполняет функцию непосредственной связи клеток животных с внешней средой, со всеми окружающими ее веществами, опорной роли не выполняет.
Под гликокаликсом животной и клеточной стенкой растительной клетки расположена плазматическая мембрана, граничащая непосредственно с цитоплазмой. В состав плазматической мембраны входят белки и липиды. Они расположены упорядоченно за счет различных химических взаимодействий друг с другом. Молекулы липидов в плазматической мембране расположены в два ряда и образуют сплошной липидный бислой. Молекулы белков не образуют сплошного слоя, они располагаются в слое липидов, погружаясь в него на разную глубину. Молекулы белков и липидов подвижны.
Функции плазматической мембраны:
– образует барьер, отграничивающий внутреннее содержимое клетки от внешней среды;
– обеспечивает транспорт веществ;
– обеспечивает связь между клетками в тканях многоклеточных организмов.
Поступление веществ в клетку
Поверхность клетки не сплошная. В цитоплазматической мембране есть многочисленные мельчайшие отверстия – поры, через которые с помощью или без помощи специальных белков, внутрь клетки могут проникать ионы и мелкие молекулы. Кроме того, некоторые ионы и мелкие молекулы могут попадать в клетку непосредственно через мембрану. Поступление важнейших ионов и молекул в клетку не пассивная диффузия, а активный транспорт, требующий затрат энергии. Транспорт веществ носит избирательный характер. Избирательная проницаемость клеточной мембраны носит название полупроницаемости .
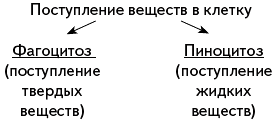
Путем фагоцитоза внутрь клетки поступают: крупные молекулы органических веществ, например белков, полисахаридов, частицы пищи, бактерии. Фагоцитоз осуществляется с участием плазматической мембраны. В том месте, где поверхность клетки соприкасается с частицей какого-либо плотного вещества, мембрана прогибается, образует углубление и окружает частицу, которая в «мембранной капсуле» погружается внутрь клетки. Образуется пищеварительная вакуоль, и в ней перевариваются поступившие в клетку органические вещества.
Путем фагоцитоза питаются амебы, инфузории, лейкоциты животных и человека. Лейкоциты поглощают бактерии, а также разнообразные твердые частицы, случайно попавшие в организм, защищая его таким образом от болезнетворных бактерий. Клеточная стенка растений, бактерий и синезеленых водорослей препятствует фагоцитозу, и потому этот путь поступления веществ в клетку у них не реализуется.
Через плазматическую мембрану в клетку проникают и капли жидкости, содержащие в растворенном и взвешенном состоянии разнообразные вещества.Это явление было названо пиноцитозом . Процесс поглощения жидкости сходен с фагоцитозом. Капля жидкости погружается в цитоплазму в «мембранной упаковке». Органические вещества, попавшие в клетку вместе с водой, начинают перевариваться под влиянием ферментов, содержащихся в цитоплазме. Пиноцитоз широко распространен в природе и осуществляется клетками всех животных.
III. Закрепление изученного материала
На какие две большие группы разделяются все организмы по строению ядра?
Какие органоиды свойственны только растительным клеткам?
Какие органоиды свойственны только животным клеткам?
Чем различается строение оболочки клеток растений и животных?
Каковы два способа поступления веществ в клетку?
Каково значение фагоцитоза для животных?
Клетка прокариот устроена значительно проще клеток животных и растений. Снаружи она покрыта клеточной стенкой, выполняющей защитные, формирующие и транспортные функции. Жёсткость клеточной стенки обеспечивает муреин. Иногда бактериальная клетка покрыта сверху капсулой или слизистым слоем.
Протоплазма бактерий, как и у эукариот, окружена плазматической мембраной . В мешковидных, трубчатых или пластинчатых впячиваниях мембраны находятся мезосомы, участвующие в процессе дыхания, бактериохлорофилл и другие пигменты. Генетический материал прокариот не образует ядра, а находится непосредственно в цитоплазме. ДНК бактерий – одиночные кольцевые молекулы, каждая из которых состоит из тысяч и миллионов пар нуклеотидов. Геном бактериальной клетки намного проше, чем у клеток более развитых существ: в среднем ДНК бактерий содержит несколько тысяч генов.
В прокариотических клетках отсутствует эндоплазматическая сеть , а рибосомы свободно плавают в цитоплазме. Нет у прокариот и митохондрий ; частично их функции выполняет клеточная мембрана.
Прокариоты
Бактерии – мельчайшие из организмов, обладающих клеточным строением; их размеры составляют от 0,1 до 10 мкм. На обычной типографской точке можно разместить сотни тысяч бактерий среднего размера. Бактерии можно увидеть только в микроскоп, поэтому их называют микроорганизмами или микробами; микроорганизмы изучаются микробиологией . Часть микробиологии, изучающая бактерии, называется бактериологией . Начало этой науке положил Антони ван Левенгук в XVII веке.
Бактерии – древнейшие из известных организмов. Следы жизнедеятельности бактерий и сине-зелёных водорослей (строматолиты) относятся к архею и датируются возрастом 3,5 млрд. лет.Из-за возможности обмена генами между представителями различных видов и даже родов систематизировать прокариот довольно сложно. Удовлетворительная систематика прокариот не построена до сих пор; все существующие системы являются искусственными и классифицируют бактерии по какой-либо группе признаков, не учитывая их филогенетического родства. Ранее бактерии вместе с грибами и водорослями включались в подцарство низших растений. В настоящее время бактерии выделены в отдельное надцарство прокариот. Наиболее распространённой системой классификации является система Берги , в основу которой положено строение клеточной стенки.
В конце XX века учёные обнаружили, что клетки сравнительно малоизученной группы бактерий – архебактерий – содержат р-РНК , отличные по своему строению и от р-РНК прокариот, и от р-РНК эукариот. Строение генетического аппарата архебактерий (наличие интронов и повторяющихся последовательностей, процессинг , форма рибосом ) сближает их с эукариотами; с другой стороны, архебактерии имеют и типичные признаки прокариот (отсутствие ядра в клетке, наличие жгутиков, плазмид и газовых вакуолей, размер р-РНК, азотфиксация). Наконец, архебактерии отличаются от всех остальных организмов строением клеточной стенки, типом фотосинтеза и некоторыми другими признаками. Архебактерии способны существовать в экстремальных условиях (например, в горячих источниках при температуре свыше 100 °С, в океанских глубинах при давлении 260 атм, в насыщенных солевых растворах (30 % NaCl)). Некоторые архебактерии выделяют метан, другие используют для получения энергии соединения серы.
По-видимому, архебактерии являются очень древней группой организмов; «экстремальные» возможности свидетельствуют об условиях, характерных для поверхности Земли в архейскую эру . Считается, что архебактерии наиболее близки к гипотетическим «проклеткам», породившим впоследствии всё многообразие жизни на Земле.
В последнее время стало ясно, что существуют три основных типа р-РНК , представленные, соответственно, первая – в клетках эукариот, вторая – в клетках настоящих бактерий, а также в митохондриях и хлоропластах эукариот, третья – у архебактерий. Исследования молекулярной генетики заставили по-новому взглянуть на теорию происхождения эукариот. В настоящее время считается, что на древней Земле одновременно эволюционировали три различные ветви прокариот – архебактерии, эубактерии и уркариоты , характеризовавшиеся разным строением и различными способами получения энергии. Уркариоты, являвшиеся, по сути, ядерно-цитоплазматическим компонентом эукариот, впоследствии включили в себя в качестве симбионтов представителей различных групп эубактерий, которые превратились в митохондрии и хлоропласты будущих клеток эукариот.
Таким образом, ранг класса, выделявшийся ранее для архебактерий, явно недостаточен. В настоящее время многие исследователи склонны разделять прокариот на два царства: архебактерии и настоящие бактерии (эубактерии ) или даже вовсе выделять архебактерии в отдельное надцарство Archaea.
Классификация настоящих бактерий приведена на схеме .
В бактериальной клетке отсутствует ядро, хромосомы свободно располагаются в цитоплазме. Кроме того, в клетке бактерии отсутствуют мембранные органоиды: митохондрии , ЭПС , аппарат Гольджи и пр. Снаружи клеточная мембрана покрыта клеточной стенкой.
Большинство бактерий передвигаются пассивно, с помощью водных или воздушных течений. Только некоторые из них имеют органеллы движения – жгутики . Жгутики прокариот очень просты по устройству и состоят из белка флагеллина, образующего полый цилиндр диаметром 10–20 нм. Они ввинчиваются в среду, продвигая клетку вперёд. По-видимому, это единственная известная в природе структура, использующая принцип колеса.
По своей форме бактерии делятся на несколько групп:
кокки (имеют округлую форму);
бациллы (имеют палочковидную форму);
спириллы (имеют форму спирали);
вибрионы (имеют форму запятой).
По способу дыхания бактерии делятся на аэробов (большинство бактерий) и анаэробов (возбудители столбняка, ботулизма, газовой гангрены). Первым для дыхания нужен кислород, для вторых кислород бесполезен или даже ядовит.
Бактерии размножаются путем деления примерно каждые 20 минут (в благоприятных условиях). ДНК реплицируется, каждая дочерняя клетка получает по своей копии родительской ДНК. Возможна также передача ДНК между неделящимися клетками (посредством захвата «голой» ДНК, при помощи бактериофагов или путём конъюгации , когда бактерии соединяются между собой копуляционными фимбриями), однако увеличения количества особей при этом не происходит. Размножению препятствуют солнечные лучи и продукты их собственной жизнедеятельности.Поведение бактерий не отличается особой сложностью. Химические рецепторы регистрируют изменения кислотности среды и концентрацию различных веществ: сахаров, аминокислот, кислорода. Многие бактерии реагируют на изменения температуры или освещенности, некоторые бактерии могут чувствовать магнитное поле Земли.
При неблагоприятных условиях бактерия покрывается плотной оболочкой, цитоплазма обезвоживается, жизнедеятельность почти прекращается. В таком состоянии споры бактерии могут часами находиться в глубоком вакууме, переносить температуру от –240 °С до +100 °С.










 – прекрасная Сунна нашего Пророка (мир ему и благословение)")
")






